Меню сайта
- На главную
- Рост растений
- Почвенные простейшие
- Полиморфизмы
- Дыхание бактерий
- Органическая природа
- Первая жизньу
- Материалы по биологии
Последние новости
Причина отравления вод океана.
Американские ученые из штата Мичиган полагают, что в качестве главной причины отравления вод Мирового океана ртутью являются бактерии.
Секрет выживания лягушек.
Американским ученым удалось выяснить, как лягушкам удается продолжать жить даже после глубокой заморозки.
Секрет долголетия ночницы.
Биологи уже давно считают, что продолжительность жизни животного определяется очень просто: чем оно больше, тем дольше живет.
Выбор физической модели кинетики энергетического метаболизмаМатериалы по биологии и химии / Исследование изменений метаболизма эритроцитов донорской крови после взятия ее из организма / Выбор физической модели кинетики энергетического метаболизмаСтраница 4
V+ = V- ≡ Vобмена (Vобмена - скорость обмена параметра).
Скорость обмена является важнейшей характеристикой проточного равновесия противоположно направленных процессов клеточного метаболизма в состоянии покоя.
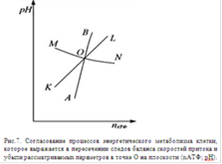
Как видно из рис. 7 кривые АВ и КL монотонно возрастают, а МN спадает, при этом все кривые пересекаются в точке О. Такая конфигурация и взаимное расположение кривых определяют устойчивость точки О по всем трем параметрам.
Рассмотрим устойчивость точки О по концентрации АТФ. В области над кривой АВ преобладают скорость V+ синтеза АТФ, а под кривой скорость V- его трат, следовательно, частная производная Vобмена концентрации АТФ по nАТФ отрицательна. При отклонении состояния метаболизма от точки О в сторону меньших концентраций АТФ, то, начнут преобладать процессы синтеза АТФ и состояние вернется в исходное устойчивое положение. То есть концентрация АТФ возрастает в результате интенсификации окислении субстратов и цитоплазма подкисляется.
Можно показать также, что точка О устойчива по рН согласно условию (1). Как говорилось, кривая МN монотонно спадает. Над кривой баланса MN преобладает скорость притока V+ ионов водорода Н+ в цитоплазму, а под кривой - скорость V- убыли Н+, значит, частная производная Vобмена ионов Н+ по рН отрицательна. При отклонении состояния метаболизма от точки О вверх по рН будут преобладать процессы подкисляющие цитоплазму, в результате которых значение рН возвращается к исходному.
Проведя аналогичные рассуждения можно заключить, что точка О также устойчива по количеству липидов в цитоплазматической мембране. Над кривой баланса KL преобладает скорость V+ синтеза липидов, а под кривой - скорость V- их ферментативного распада. При переходе через точку О слева направо частная производная Vобмена количества липидов по nАТФ отрицательна, также при переходе через точку О снизу вверх частная производная по рН также отрицательна. Следовательно, согласно условию (1) эта точка устойчива по количеству липидов С в цитомембране.
Для дальнейшего анализа кинетики метаболизма эритроцитов важно получить указанные следы кривых баланса скоростей V- и V+ для основных параметров, используемых в принятой за основу модели, во всем физиологическом диапазоне изменений концентрации АТФ и рН, включая область больших, чем в состоянии покоя, значений концентрации АТФ. Ясно, что конфигурации и взаимное расположение кривых, обеспечивающих устойчивость новой точки 3 с координатами (nАТФ3,рН3), в случае согласованности процессов метаболизма, будут подобны рассмотренному выше случаю для состояния покоя клетки. Для эритроцитов это имеет место. Для подавляющего большинства соматических клеток (кроме возбудимых клеток) согласно работе [16] в области высоких значений nАТФ устойчивости состояний метаболизма по рН нет. Поэтому для дальнейшего анализа кинетики метаболизма эритроцитов мы воспользуемся следующей моделью, которая проиллюстрирована на рис. 8.

Кривая ABCD на фазовой плоскости (nАТФ; рН) является следом кривой баланса скоростей синтеза V+ и расхода V- АТФ. Форма этой кривой имеет зигзагообразный вид. Точки ветвей АВ и CD устойчивы по условию (1), кроме точек В и С, которые неустойчивы по условию (2). Точки ветви ВС не устойчивы по условию (1).
Из таких же соображений получена форма кривой KLК1L1. Точки ветвей участков кривой KL и К1L1 устойчивы по условию (1), а точки ветви LK1 не устойчивы по этому условию. Точки L и K1 неустойчивы по условию (2).
Для качественного анализа (достаточного для получения необходимых результатов) хода кривой баланса MN не требуется строить соответствующие поверхности. Расположение и характерную конфигурацию кривой MN можно получить аналитически. Имея начальную точку, для определения конфигурации и расположения на плоскости (nАТФ, рН) кривой MN, можно воспользоваться известной зависимостью рН цитоплазмы от величины трансмембранного потенциала.
Поскольку цитомембрана не представляет значительного препятствия для ионов Н+, то можно использовать соотношение Больцмана:
![]() (3),
(3),
где ![]() - концентрация ионов
- концентрация ионов ![]() в цитоплазме эритроцитов;
в цитоплазме эритроцитов; ![]() - концентрация ионов
- концентрация ионов ![]() в плазме крови;
в плазме крови; ![]() - элементарный заряд;
- элементарный заряд; ![]() - трансмембранный потенциал; k - постоянная Больцмана; T - температура. Прологарифмировав соотношение (1), получим зависимость рН цитоплазмы от концентрации АТФ(4):
- трансмембранный потенциал; k - постоянная Больцмана; T - температура. Прологарифмировав соотношение (1), получим зависимость рН цитоплазмы от концентрации АТФ(4):
Смотрите также
Морфология и анатомия бактериальных организмов
«Мир
беспорядоченно
усеян
упорядоченными формами»
Поль
Валери
...
Роль биологических мотиваций в поведении
Введение
Термин "мотивация"
имеет латинский корень movere - двигать. Зоологи, наблюдая за поведением
животных в природе, также выдвинули предположение о существовании механи ...
Первая жизнь
Введение
За
последние годы учёные обнаружили впечатляющие свидетельство, того как выглядели
первые создания на земле. Подсказки к разгадке этой тайны можно найти и среди
современных ...