Меню сайта
- На главную
- Рост растений
- Почвенные простейшие
- Полиморфизмы
- Дыхание бактерий
- Органическая природа
- Первая жизньу
- Материалы по биологии
Последние новости
Причина отравления вод океана.
Американские ученые из штата Мичиган полагают, что в качестве главной причины отравления вод Мирового океана ртутью являются бактерии.
Секрет выживания лягушек.
Американским ученым удалось выяснить, как лягушкам удается продолжать жить даже после глубокой заморозки.
Секрет долголетия ночницы.
Биологи уже давно считают, что продолжительность жизни животного определяется очень просто: чем оно больше, тем дольше живет.
Выбор физической модели кинетики энергетического метаболизмаМатериалы по биологии и химии / Исследование изменений метаболизма эритроцитов донорской крови после взятия ее из организма / Выбор физической модели кинетики энергетического метаболизмаСтраница 3
Таким образом, изменяя скорости протекания процессов синтеза и трат АТФ с помощью их активации и ингибирования, можно переводить систему из одного устойчивого состояния в другое, если данная физическая модель допускает существование двух устойчивых состояний энергетического метаболизма эритроцита.
В свете изложенного выше, взаимные переходы между состояниями эритроцита с высокой и низкой концентрацией АТФ осуществляются тогда, когда кривые V+ синтеза и V- расхода АТФ смещаются в противоположные стороны (сближаются либо расходятся). Это смещение может достигаться за счет ингибирования процесса синтеза АТФ и одновременной активации процесса его трат. На основе анализа кинетики метаболизма эритроцитов с помощью выбранной за основу физической модели нельзя утверждать, что всегда существует вторая устойчивая точка, лежащая в области низких значений АТФ, поскольку эта модель допускает варианты, когда такой точки не существует. Для данного исследования интересны не все возможные ситуации, а только те, когда энергетический метаболизм эритроцитов близок к in vivo. Для прояснения вопроса о том, всегда ли существует состояние эритроцитов с низкой nАТФ, требуется привлечение более точной модели, которая возможно даст однозначный ответ о наличии в этих условиях указанной устойчивой точки.
Рассмотрим модель равновесных процессов метаболизма, предложенную Фоком М.В. и соавторами в работе [10], дополнив ее некоторыми деталями и назвав обобщенной моделью, как это сделано в работе [15]. В этой работе [15] рассматривается состояние покоя клеток, режимы их активности и деления. Нас интересует только состояние покоя, в котором достигается гомеостаз всех параметров, обеспечивающий жизнеспособность и жизнедеятельность клеток.
В обобщенной модели основными параметрами при анализе кинетики энергетического метаболизма выбраны показатель кислотности рН цитоплазмы, концентрация АТФ в ней и количество липидов C в мембране. Первые два параметра относятся к быстроменяющимся параметрам, так как величины их могут изменяться за доли секунды, то есть значительно быстрее времени обращения крови по большому кругу. В отличие от этих параметров, количество липидов в мембране эритроцитов меняется медленно, но также является важным энергетическим параметром, который определяет жизнеспособность и целостность клетки. Количество ферментов, запасы глюкозы в цитоплазме, объем эритроцитов и др. в модели приняты постоянными. Свободными же оставляют 2 переменные - nАТФ и рН цитоплазмы, которые определяют скорости таких энергозависимых процессов в эритроцитах как скорости синтеза и распада АТФ, скорости подкисления и подщелачивания цитоплазмы, а также скорости синтеза и распада липидов. Эти скорости будут функциями только двух указанных переменных.
![]() ,
,![]() - скорости синтеза и расхода АТФ;
- скорости синтеза и расхода АТФ;
![]() ,
,![]() - скорости подкисления подщелачивания цитоплазмы;
- скорости подкисления подщелачивания цитоплазмы;
![]() ,
,![]() - скорости синтеза и распада липидов.
- скорости синтеза и распада липидов.
Зависимости скоростей каждого из перечисленных процессов от рН и nАТФ в трехмерном пространстве величин (pH, nАТФ, V - скорости процессов) могут быть представлены сложными поверхностями [15, 16]. На линиях пересечения поверхностей, полученных для каждых двух противоположно направленных процессов (синтез и расход АТФ, подкисление и подщелачивание цитоплазмы, синтез и распад липидов в мембранных структурах), достигается баланс их скоростей.
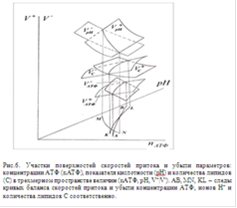
На рис. 6 представлены участки этих поверхностей. Их линии пересечения трехмерны. Пара участков поверхностей V+АТФ и V-АТФ дают линию пересечения, в точках которой скорость синтеза АТФ равна скорости расхода АТФ; пара поверхностей V+рН и V-рН дают линию пересечения, в точках которой скорость подкисления равна скорости подщелачивания цитоплазмы; пара поверхностей V+С и V-С дают линию пересечения, в каждой точке которой скорость увеличения равна скорости уменьшения количества липидов в мембране. Следами этих линий пересечения (в проекции на плоскость (nАТФ; рН)) являются кривые АВ, MN и KL, соответственно, для каждой из перечисленных выше линий пересечения. Как видно из рис. 6, эти кривые попарно пересекаются, что отражает рассогласованность процессов метаболизма.
На рис. 7. эти следы показаны на фазовой плоскости (nАТФ; рН) для случая согласования процессов метаболизма. При значениях рН и nАТФ, соответствующих точкам на этих кривых,
Смотрите также
Асептика в биотехнологии
Введение
Биотехнологические процессы в основном проводят в асептических
условиях. Асептика - это комплекс мероприятий, направленных на предотвращение
попадания в среду посторонних вещ ...
Классификация насекомых. Первичнобескрылые
ВВЕДЕНИЕ
Насеко́мые (лат. Insecta) - класс шестиногих членистоногих
(Arthropoda) животных, группа, обладающая наибольшим разнообразием среди всех
остальных животных на Земле. Изв ...
Сон и бодрствование у человека и животных
Введение
С древнейших времен, вероятно, еще с тех пор, как человек обрел сознание
и только начал мыслить, его глубоко интересует вопрос о природе и механизмах
одного из самых таинств ...