Меню сайта
- На главную
- Рост растений
- Почвенные простейшие
- Полиморфизмы
- Дыхание бактерий
- Органическая природа
- Первая жизньу
- Материалы по биологии
Последние новости
Причина отравления вод океана.
Американские ученые из штата Мичиган полагают, что в качестве главной причины отравления вод Мирового океана ртутью являются бактерии.
Секрет выживания лягушек.
Американским ученым удалось выяснить, как лягушкам удается продолжать жить даже после глубокой заморозки.
Секрет долголетия ночницы.
Биологи уже давно считают, что продолжительность жизни животного определяется очень просто: чем оно больше, тем дольше живет.
Строение и функции гемоглобинаМатериалы по биологии и химии / Исследование изменений метаболизма эритроцитов донорской крови после взятия ее из организма / Строение и функции гемоглобинаСтраница 1
Гемоглобин
- главный компонент эритроцита и основной дыхательный пигмент, обеспечивающий перенос кислорода (О2) из легких к тканям в организмах позвоночных животных, и, в меньшей степени, углекислого газа (СО2) от тканей в легкие. Главным образом, углекислота переносится в виде ![]() в плазме крови. Подсчитано, что в одном эритроците содержится ~ 340.000.000 молекул гемоглобина, каждая из которых состоит примерно из 103 атомов. В крови человека в среднем содержится ~ 750г гемоглобина [3].
в плазме крови. Подсчитано, что в одном эритроците содержится ~ 340.000.000 молекул гемоглобина, каждая из которых состоит примерно из 103 атомов. В крови человека в среднем содержится ~ 750г гемоглобина [3].
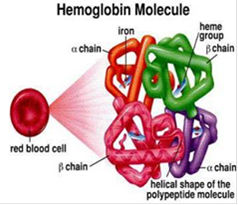
Гемоглобин представляет собой сложный белок, относящийся к группе гемопротеинов. Белковый компонент его представлен глобином, состоящим из двух a и двух b субъединиц и, соответственно, содержит четыре полипептидные цепочки двух сортов. Небелковый компонент (так называемая простетическая группа) представлен четырьмя одинаковыми железопорфириновыми соединениями - гемами.
Иржак Л.И. в своей работе [4] описал большинство физиологических свойств гемоглобина, его виды и подвиды. Основное внимание в этой работе уделено описанию процессов связывания кислорода гемоглобином в норме и при некоторых патологиях. Также приведена кривая диссоциации оксигемоглобина и описан эффект Бора.
Наиболее подробно физико-химические свойства гемоглобина изучены и представлены в работах Блюменфельда Л.А. [5, 6]. В них большое внимание уделено зарядовым характеристикам молекул гемоглобина. Эти характеристики в значительной степени определяют адгезивные свойства молекул гемоглобина (комплексование и ассоциацию с липидной мембраной). В указанных работах приведены теоретические и экспериментальные исследования, в которых сравниваются свойства окси- и деоксигемоглобина, а также гемоглобина и миоглобина.
При взаимодействии молекулярного кислорода с гемоглобином в условиях in vivo существует небольшая, но конечная вероятность окисления гема двухвалентного железом до трехвалентного, причем молекула О2 не присоединяется, но окисляет железо: Fe2+ + O2 Þ Fe3+ O2-. Поэтому при дыхании в эритроцитах непрерывно образуется метгемоглобин, который не способен переносить О2. Восстановление Fe3+ метгемоглобина до Fe2+ происходит с затратами энергии и обеспечивается ферментом - метгемоглобинредуктазой [3].
В ходе метаболизма глюкозы, протекающего в эритроците по побочному пути гликолиза, контролируемого ферментом дифосфоглицератмутазой, в эритроците образуется 2,3-дифосфоглицерат (2,3-ДФГ). Основное его значение заключается в уменьшении сродства гемоглобина к кислороду. 2,3-ДФГ связывается с β-цепями гемоглобина, облегчая отсоединение О2 от молекулы гемоглобина [3].
Большое количество опубликованных работ посвящено строению мембран клеток, в том числе и эритроцитов. В одной из классических работ [7] приведены данные по количеству и белковому составу мембран эритроцитов и функциональному назначению этих белков, архитектонике липидного бислоя.
Методики и результаты идентификации некоторых видов белков мембранных структур представлены в работе [8]. В мембране эритроцита идентифицировано 15 главных белков. Более 60 % всех белков составляют: примембранный белок спектрин и мембранные белки - гликофорин и белок полосы 3.
Спектрин является белком цитоскелета, связанным с внутренней стороной мембраны. Он участвует в поддержании двояковогнутой формы эритроцита.
Гликофорин - белок, относящийся к классу мембранных гликопротеинов, по мнению авторов работ [9, 10, 11], играет важную роль в механизме авторегуляции неспецефической проницаемости цитоплазматической мембраны эритроцита. Этот белок пронизывает мембрану в виде одиночной спирали. Большая его часть выступает с наружной поверхности эритроцита, где к нему присоединены 15 отдельных цепей олигосахаридов, на ветвях которых расположены сиаловые кислоты, несущие отрицательные заряды. В распрямленном виде максимальное удаление ветвей гликофорина от поверхности эритроцита может достигать 200 нм, что больше поляризационной длины плазмы крови (радиуса экранировки Дебая). В результате этого отдельные отрицательные заряды экранированы противоионами плазмы, так что нити гликофорина не взаимодействует с поверхностью своей мембраны. Так как противоионы могут быть общими для гликофоринов различных эритроцитов, то эритроциты могут образовывать монетные столбики.
Смотрите также
Племенное разведение немецкой овчарки
Введение
Немецкая овчарка - одна из самых любимых собачьих пород. Имеет довольно
широкое применение: может служить собакой-компаньоном, охранной, защитной,
сыскной, служебной и ...
Классификация насекомых. Первичнобескрылые
ВВЕДЕНИЕ
Насеко́мые (лат. Insecta) - класс шестиногих членистоногих
(Arthropoda) животных, группа, обладающая наибольшим разнообразием среди всех
остальных животных на Земле. Изв ...
Клебсиеллы. Патогенность
Введение
Род Klebsiella, названный так в честь Э. Клебса, включает 4 вида, первый из
которых Klebsiella pneumoniae состоит из трех подвидов: K. subsp. pneumoniae, K. ozaenae и K. rhino ...